


Annual survival and population cycles
Annual survival
 Most black grouse mortaility is caused by predation. Depending on the region, this can be mainly from raptors such as goshawks Accipter gentilis, but also mammalian predators such as foxes Vulpes vulpes and mustelids (martens Martes spp. or stoats Mustela spp.). Annual survival varies between population. It can be as high as 0.72 (adults) from England (Warren & Baines 2002). Typically, survival rates are lower in Scandinavia e.g.
0.53 (Finland: Lindén 1981) and 0.54 (Sweden: Angelstam 1984). In Central
Europe and the French Alps estimates of survival rates of
adults are higher 0.66 survival (hens only) in the
Netherlands (Vos 1983) and 0.56-0.84 in the French Alps (Caizergues
& Ellison 1997). However, there can be considerable variation in survival rates between years e.g. 0.28-0.52 in Scandinavia (Willebrand 1988).
Most black grouse mortaility is caused by predation. Depending on the region, this can be mainly from raptors such as goshawks Accipter gentilis, but also mammalian predators such as foxes Vulpes vulpes and mustelids (martens Martes spp. or stoats Mustela spp.). Annual survival varies between population. It can be as high as 0.72 (adults) from England (Warren & Baines 2002). Typically, survival rates are lower in Scandinavia e.g.
0.53 (Finland: Lindén 1981) and 0.54 (Sweden: Angelstam 1984). In Central
Europe and the French Alps estimates of survival rates of
adults are higher 0.66 survival (hens only) in the
Netherlands (Vos 1983) and 0.56-0.84 in the French Alps (Caizergues
& Ellison 1997). However, there can be considerable variation in survival rates between years e.g. 0.28-0.52 in Scandinavia (Willebrand 1988).
Population cycles
Some black grouse populations, particularly those in boreal forests, undergo cyclical changes in population density. The changes are both regular, occur in synchrony with other species (other forest grouse such as capercaillie Tetrao urogallus) and occur of large spatial scales. These fluctuation occur in 6-7 year cycles and hunting bag data in Finland suggests they have been present since at least the early 1900's (Lindström et al. 1995; Ranta et al. 1995). This cyclicity seems also strongly connected to the large-scale synchrony. Local populations commonly fluctuate in synchrony, and the degree of synchrony declines with distance between the populations. Likewise, grouse populations of different species in the same area commonly fluctuate in synchrony as well.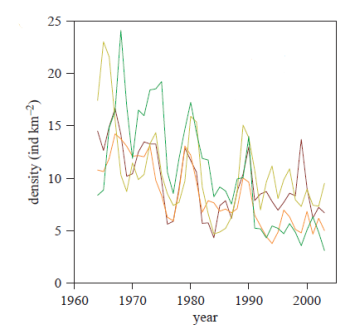 Interestingly, both the cycles and the synchrony have apparently disappeared by the middle of the 1980s (Ranta et al. 2004).
Interestingly, both the cycles and the synchrony have apparently disappeared by the middle of the 1980s (Ranta et al. 2004).
References
Angelstam, P. (1984) Sexual and seasonal differences in mortality of the Black Grouse Tetrao tetrix in boreal Sweden. Ornis Scandinavica, 15. 123-124.
Caizergues, A. & Ellison, L.N. (1997) Survival of Black Grouse Tetrao tetrix in the French Alps. Wildlife Biology, 3, 177- 188.
Lindén, H. (1981) Estimation of juvenile mortality in the Capercaillie Tetrao urogallus and the Black Grouse T. tetrix from indirect evidence. Finnish Game Research, 39, 35-51.
Lindström, J., Ranta, E., Kaitala, V. & Lindén, H. (1995) The clockwork of Finnish tetraonid population dynamics. Oikos 74, 185-194.
Ranta, E., Lindström J. and Lindeacute;n H. (1995) Synchrony in tetraonid population dynamics. Journal of Animal Ecology, 64, 767-776.
Ranta, E., Helle, P. & Lindeacute;n, H. (2004) Forty years of grouse monitoring in Finland. Suomen Riista, 50, 128-136 (in Finnish with English summary).
Vos, G.S. (1983) Social behavior of black grouse: an observational study. Ardea, 71, 1-103.
Warren, P.K. & Baines, D. (2002) Dispersal, survival and causes of mortality in black grouse Tetrao tetrix in northern England. Wildlife Biology, 8, 129–135.
Willebrand, T. (1988) Demography and ecology of Black Grouse populations. PhD thesis, University of Uppsala, Sweden, 148 pp.